植物激素生长素通过下游信号通路调控植物的生长发育,包括广为人知的SCFTIR1/AFB-Aux/IAA-ARF通路和其他几种不太典型的“非经典”通路。最近,通过分析生长素在拟南芥顶端钩发育中的功能,发现了一种由Transmembrane Kinase 1(TMK1)介导的生长素非经典信号途径(Cao et al. 2019)。生长素在顶端钩凹侧的不对称积累触发DAR1催化的TMK1 C末端的释放(Gu et al. 2022),TMK1c入核后磷酸化并稳定IAA32/34以抑制细胞伸长,这对黄化苗顶钩的形态建成至关重要。
4月18日,金沙集团1862cc成色、教育部细胞活动与逆境适应重点实验室、甘肃省基因编辑育种重点实验室的郭光沁团队在PNAS上发表论文,报道了CYTOKININ INDUCED ROOT WAVING 1 (CKRW1) / WAVY GROWTH 3(WAV3)亚家族蛋白作为E3泛素连接酶参与TMK1介导的生长素非典型信号调控的顶钩形态建成。该论文第一作者为金沙集团1862cc成色汪军丽博士,通讯作者为郭光沁教授和青年教师武磊。
原文链接:https://www.pnas.org/doi/10.1073/pnas.2314353121

研究者首先证明了WAV3亚家族蛋白具有E3泛素连接酶活性,后续通过构建缺乏酶活的突变型蛋白WAV3H147Y和WAV3C125S,发现其泛素连接酶活性是顶钩发育所必需的(图1)。紧接着找到了并验证了WAVE3泛素连接酶的底物蛋白是AUX/IAA蛋白IAA32和IAA34,并且验证了发生泛素化修饰的IAA32和IAA34通过26S蛋白酶体途径降解(图2)。
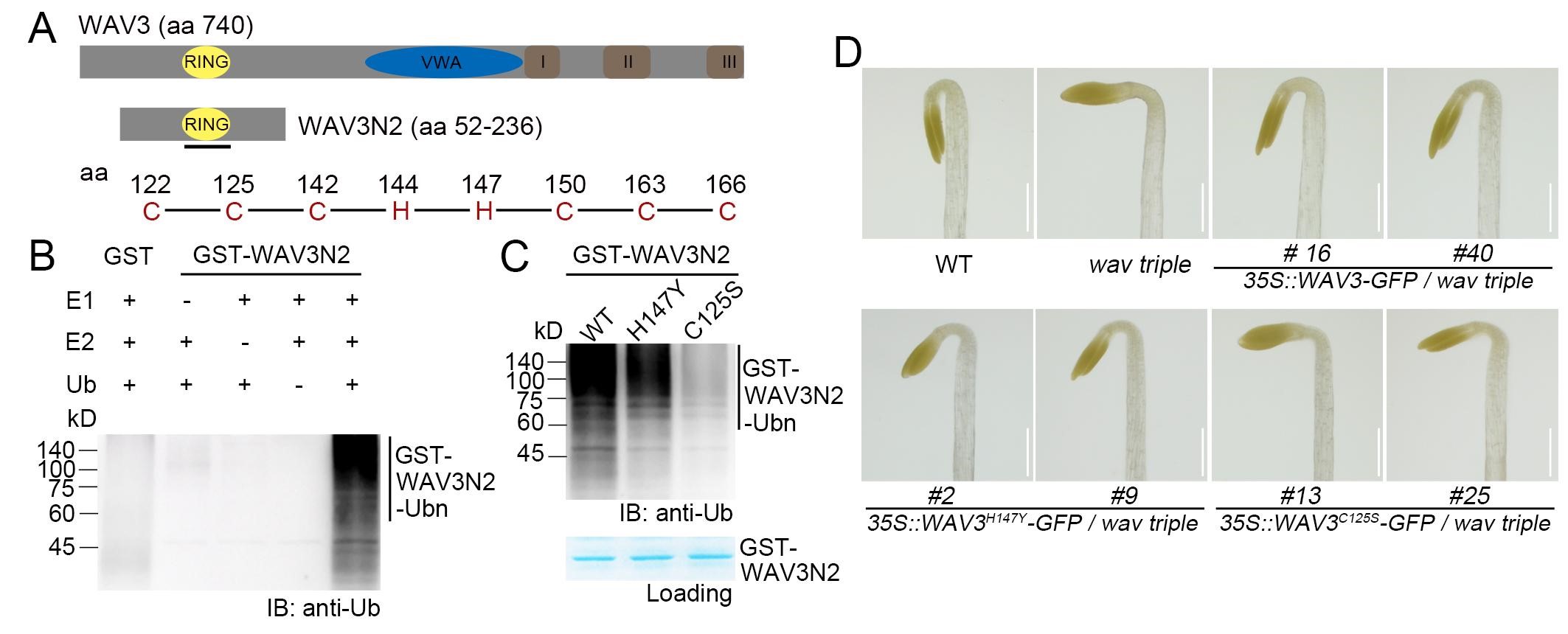
图1 WAV3具有泛素连接酶活性,且该酶活性与顶钩的形态建成相关

图2 WAV3靶向IAA32/34泛素化,泛素化修饰促进IAA32/34的降解。
研究者通过观察融合有GFP荧光标记的IAA32/34-GFP在WAV三突变背景下(wav triple)的分布,发现IAA32和IAA34在wav triple的顶钩凸面异常积累,并且这种积累导致了wav triple顶钩凸面细胞的伸长被抑制,这也是该突变体顶钩形态建成缺陷的原因(图3)。有趣的是WAV E3泛素连接酶并不降解顶钩凹面的IAA32和IAA34。研究者结合之前Caoetal. 2009中TMK1c介导的磷酸化可以稳定IAA32和IAA34的结论。通过实验证明了TMK1c和WAV E3泛素连接酶之间相互拮抗,共同调节暗形态建成中顶钩凹面和凸面细胞中IAA32和IAA34的不对称分布,从而决定两面细胞差异化伸长(图4)。该过程的模型如图5所示。
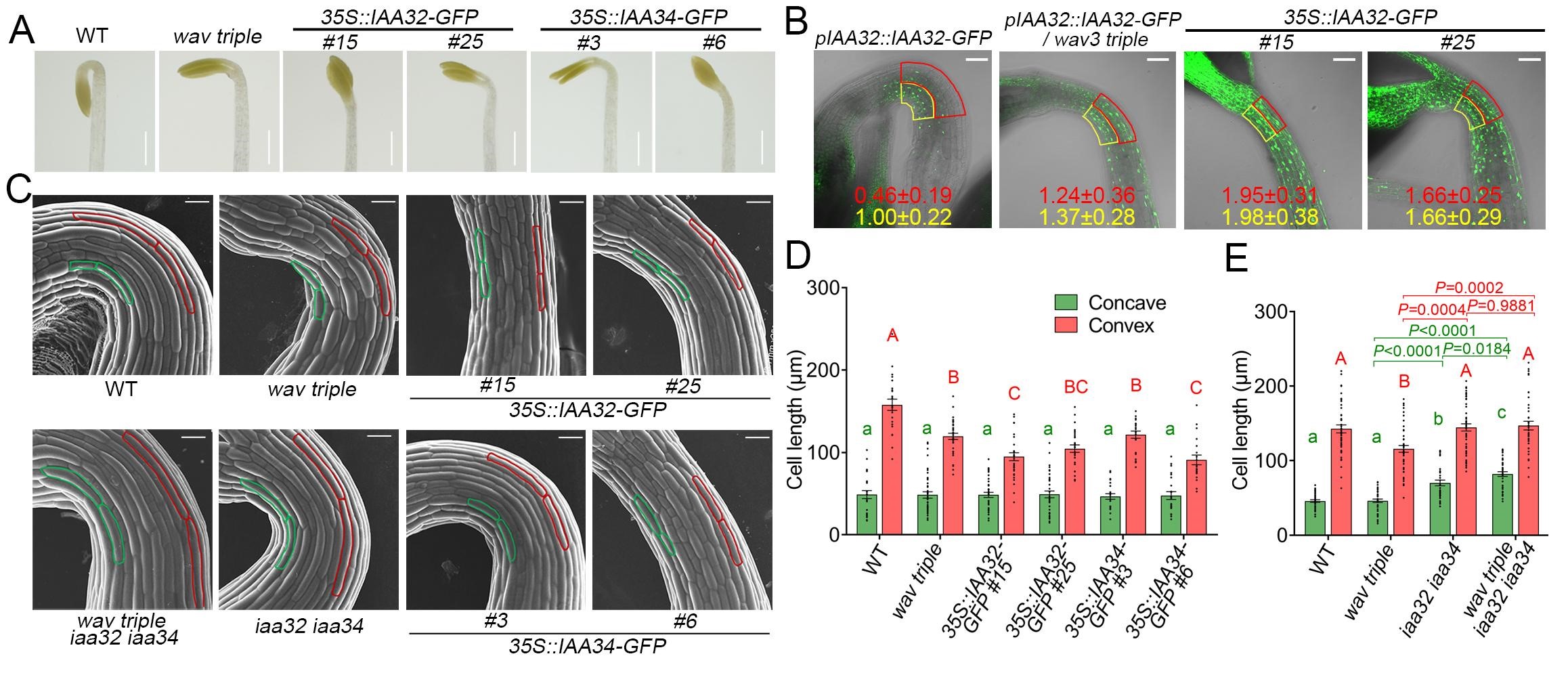
图3 WAV E3泛素连接酶介导顶钩凸面IAA32和IAA34的降解。
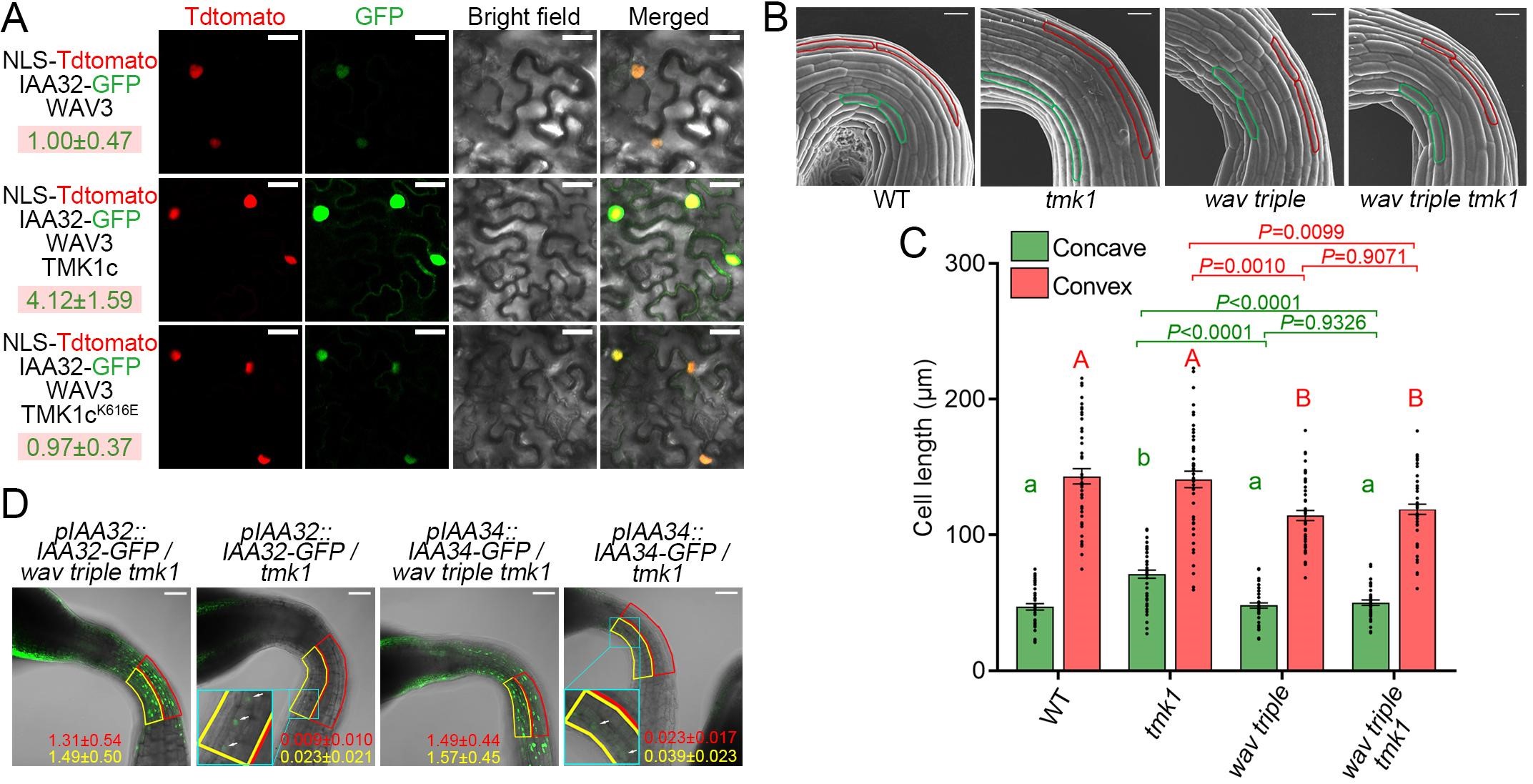
图4 在顶钩凹面TMK1c通过磷酸化阻止WAV E3介导IAA32/34降解。
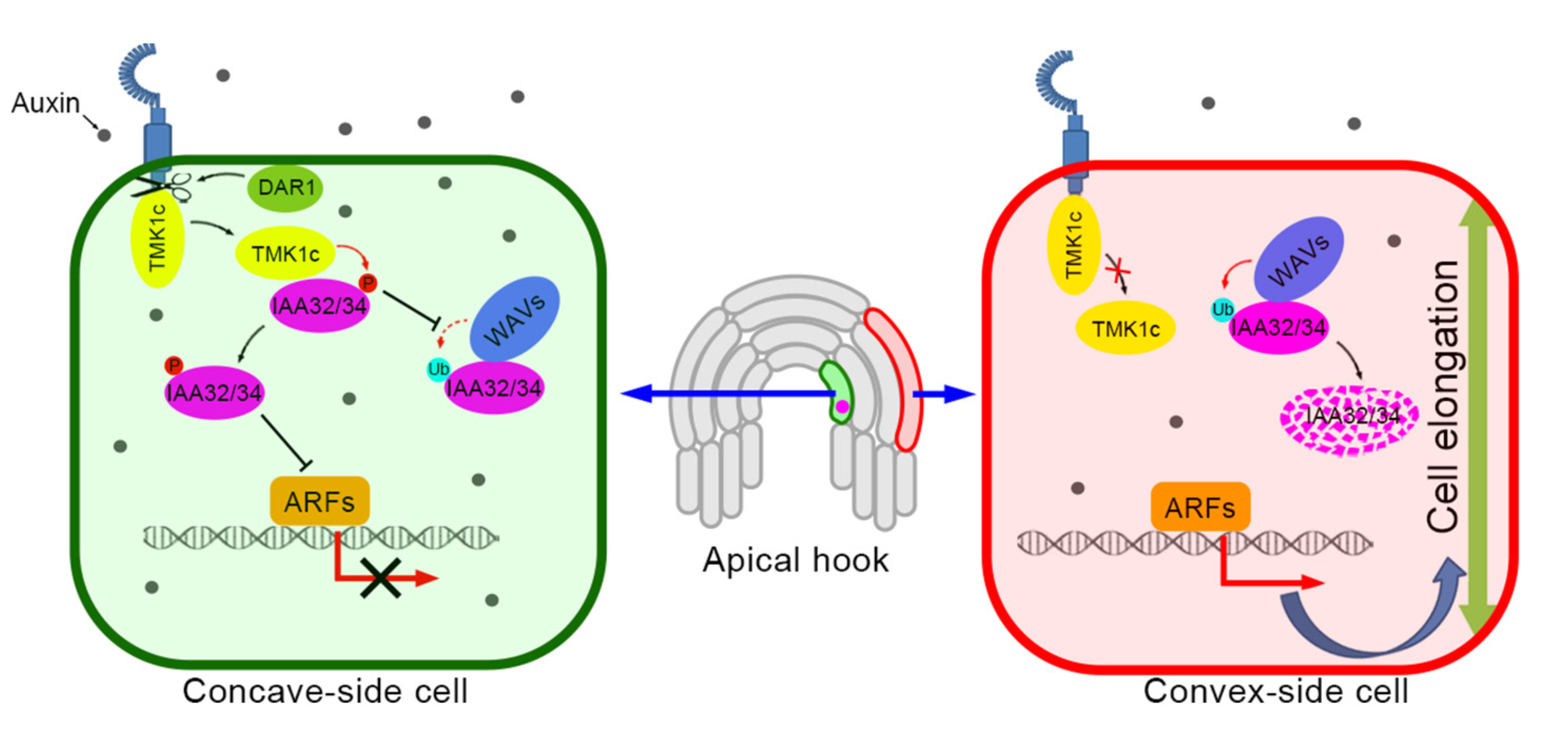
图5 WAV和TMK1共同调控顶钩的形态建成的模型。
近20年来,郭光沁团队一直连续系统地从事生长素突变体的分离和研究工作:1)是国际上最早独立发现IAA合成酶基因CKRC1/TAA1的实验室之一(Zhou et al. 2011);2)随后通过大规模筛选,获得了一批新的生长素缺陷突变体(Wu et al. 2015);通过对这些突变体的研究,3)揭示了细胞分裂素调控CKRC1/TAA1和CKRC2/YUC8基因转录和生长素合成的一条分子途径(CK-ARR1/12-PIF4途径)(Di et al. 2016);4)发现CKRW2/HUB1和HUB2复合体介导的H2B单泛素化修饰促进部分IAA合成基因的表达和生长素合成(Zhang et al. 2021);5)发现由CKRC3/TCU2/NAA25和NBC/NAA20组成的NatB复合体通过介导YUC/ACOs蛋白NTA修饰,维持蛋白稳定性和植物体内IAA/乙烯稳态(Liu et al. 2021; 2022);6)发现CKRW1/WAV3亚家族E3泛素连接酶介导非典型IAA32和IAA34的泛素化和降解,参与生长素对顶勾发育的调控(Wang et al. 2024)。上述研究得到了国家自然科学基金委、教育部、甘肃省科技厅等机构的资助。


